Altern: Chancen und Risiken
Von Beginn an begleitet den Menschen das Erleben seiner Endlichkeit bzw. der frühe Tod innerhalb der Familie. Über lange Zeit hinweg (seit ca. 50.000 Jahren) lag die mittlere Lebenserwartung zwischen 30 und 40 Jahren.1 Diese an sich kurze Lebensspanne war den Menschen wohl bewusst. So sagte bereits der römische Philosoph und Politiker Lucius Annaeus Seneca (ca. 4 v. bis 65 n. Chr.): »Bemiss deine Lebenszeit, für so vieles reicht sie nicht«. Auf der anderen Seite wurde schon immer versucht, diese kurze Lebensspanne positiv zu beeinflussen. Leonardo da Vinci (1452 bis 1519) hatte bereits früh erkannt, dass dazu eine lebenslange Vorbereitung gehört: »Was man in seiner Jugend erwirbt, dient im Kampf gegen das Elend des hohen Alters. Und wenn du willst, dass dein Alter sich aus Weisheit nähre, so sorge dafür, solange du jung bist, dass es in deinem Alter nicht an Nahrung mangelt« (Codex Atlanticus). Trotz der nach heutigen Lebensumständen niedrig erscheinenden damaligen mittleren Lebenserwartung wird die entsprechende maximale Lebenserwartung im selben Zeitraum immerhin auf knapp unter 100 Jahren geschätzt.2 Damit hat es immer auch Menschen gegeben, die sehr alt geworden sind. Da diese alten Menschen zu ihrer Zeit aber eine große Ausnahme waren und zum Teil sogar zwei Generationen ›überlebt‹ hatten, wurden sie geehrt und für ihr wichtiges Erfahrungswissen geschätzt. Interessanterweise sind die meisten historischen Berichte über die Lebenserwartung bzw. das Alter der Menschen geschlechtsbezogen verzerrt. So gibt es wesentlich mehr Daten zu Männern als zu Frauen, da die Männer oft als das wichtigere Geschlecht angesehen wurden. Dies wird indirekt auch bei den Darstellungen des Stufenalters der Menschen aus dem 18. und 19. Jahrhundert sichtbar. Dort wird zumeist der Mann in allen seinen Lebenslagen abgebildet, die Frau indes begleitet ihn nur in jungen Jahren als Teil der Familie mit den Kindern (Abb. 1).
 Abb. 1: Das Stufenalter des Menschen. Kolorierte Lithographie (um 1855). Neuruppiner Bilderbogen Nr. 2740. Foto: bpk / Museum Europäischer Kulturen, SMB.
Abb. 1: Das Stufenalter des Menschen. Kolorierte Lithographie (um 1855). Neuruppiner Bilderbogen Nr. 2740. Foto: bpk / Museum Europäischer Kulturen, SMB.
Die Lebenserwartung hat sich in den letzten 150 Jahren nahezu verdoppelt. Dieser lineare Anstieg wurde von Jim Oeppen und James W. Vaupel 2002 für die Frauen weltweit beschrieben.3 Interessanterweise hatten 1840 die Skandinavierinnen die höchste Lebenserwartung, wohingegen diese nun bei den Japanerinnen am höchsten ist. Während zu Beginn vor allem Verbesserungen von Hygiene, Kindersterblichkeitsrate, Behandlungsmöglichkeiten von Infektionserkrankungen und Ernährung ausschlaggebend für die Erhöhung der Lebenserwartung waren, ist es jetzt vor allem die bessere medizinische Versorgung chronischer Erkrankungen bei alten Patienten. Das höchste bisher nachgewiesene Lebensalter beim Menschen erreichte Jeanne Calment (21.2.1875 bis 4.8.1997) mit 122 Jahren. Berichte über Menschen mit noch höherem Alter konnten bisher nicht verifiziert werden, da in den meisten Fällen der Nachweis der Geburt sehr schwierig ist. Ob wie bisher die mittlere Lebenserwartung weiter so stark ansteigen wird, wird erst die Zukunft zeigen, denn die Zunahme von Menschen mit dem metabolischen Syndrom oder Diabetes könnten diesen linearen Anstieg auch unterbrechen.4 Für die Gesellschaft per se ist der Anstieg der Lebenserwartung eher unproblematisch. Viel problematischer ist dagegen die gesunkene Geburtenziffer (Anzahl der Kinder pro gebärfähige Frau) zu bewerten. Diese liegt in Deutschland seit Beginn der 1970er Jahre weit unter zwei, also unter dem Wert, den man bräuchte, um eine Population stabil zu halten. Parallel dazu kam es in den Jahren nach der Wiedervereinigung beider deutschen Staaten zu einer großen Bevölkerungswanderung; überwiegend junge Menschen in den neuen Bundesländern sind aufgrund der wesentlich besseren Lebensperspektiven in die alten Bundesländer abgewandert. Das hat das mittlere Alter in den neuen Bundesländern stark steigen lassen. Parallel dazu gibt es nun den wachsenden Trend der Verstädterung. Da die Versorgung im ländlichen Gebiet immer schwieriger wird, ziehen viele Menschen in die Städte, was den »Alterungsprozess« der Gesellschaft auf dem Lande weiter beschleunigt.
Doch was ist eigentlich alt und wie altern wir Menschen? Die Weltgesundheitsorganisation (WHO) hat sich bei der Definition »alter Mensch« nach dem Rentenantrittsalter gerichtet. Demnach ist ein Mensch alt, wenn er das 65. Lebensjahr erreicht hat. Allerdings gibt es den alten Menschen so nicht, denn es gibt sowohl fitte 80-Jährige als auch verbrauchte 60-Jährige. Damit sagt das kalendarische Alter also wenig über das tatsächliche biofunktionale Alter aus. Dies wird insbesondere klar, wenn man sich Fotos verschiedener 60-jähriger Menschen anschaut, deren optische ›Altersunterschiede‹ frappant sein können. Neben den rein optischen Unterschieden gibt es zudem viele messbare physiologische Parameter wie z. B. den Pulsdruck, die die große Heterogenität im Alter nachvollziehbar machen.5
Das Altern kann biomedizinisch als eine Anreicherung molekularer Schäden begriffen werden, wobei die Entstehung dieser Schäden die treibende Kraft des Alterns darstellt und die Reparaturfähigkeit die Lebenserwartung widerspiegelt.6 Die Anreicherung molekularer Schäden geschieht ein Leben lang und beginnt bereits mit der Geburt. Dies wird im Allgemeinen während der Kindheit und im jungen Erwachsenenalter noch nicht gesehen, da man keinen Phänotyp entwickelt. Molekulare Schäden können prinzipiell an allen Makromolekülen entstehen, d. h. an den Nukleinsäuren, den Fetten sowie den Proteinen. Aus biologischer Sicht sind mehrere Mechanismen beim Altern ursächlich beteiligt. Dazu gehören der oxidative Stress mit den Radikalen, der glukogene Stress mit der Glykierung, die Inflammation, die Reduktion der Telomerlänge, veränderte hormonelle Verhältnisse, reduzierte regenerative Kapazitäten und viele weitere mehr.7 Kein einziger dieser Faktoren kann den Alterungsprozess komplett erklären, denn es sind immer viele Faktoren, die im Zusammenspiel für diesen Prozess verantwortlich sind. Des Weiteren ist das zu betrachtende Zielorgan für die Einschätzung der Bedeutung verschiedener Alterungsprozesse von großer Bedeutung. So ist für ein Gewebe mit vielen proliferierenden (sich neu bildenden) Zellen ein anderes Zusammenspiel an Parametern (z. B. zelluläre Seneszenz, Telomerlänge) wichtiger als bei Geweben mit wenig proliferierenden Zellen (z. B. Oxidation, Glykierung). Zu den Geweben mit hohen Proliferationsraten zählen z. B. Haut, Darm und Blut, zu denen mit geringen Proliferationsraten z. B. Gehirn und Herz. Für den Menschen sind aber gerade die Organe, die wenige proliferationskompetente Zellen und somit geringe Regenerationsfähigkeit haben, im Alter wichtig. So sind es die großen degenerativen Erkrankungen wie die Demenz oder die Herzinsuffizienz, die das Leben im Alter entscheidend prägen können. Daher wird den proliferationsunabhängigen Alterungsprozessen eine wichtige Rolle zugesprochen.
Zu den wichtigen proliferationsunabhängigen Alterungsprozessen gehört die Glykierung als eine chemische Reaktion, die keinerlei Enzyme benötigt. Diese Reaktion wurde 1912 erstmalig für Proteine bzw. Aminosäuren von Louis Camille Maillard (1878 bis 1936) als Bräunungsreaktion beschrieben.8 Die Produkte dieser Reaktion werden fortgeschrittene Glykierungsendprodukte oder »Advanced Glycation Endproducts« (AGEs) genannt. In der klassischen Maillardreaktion kommt es zu einer Addition von Monosacchariden wie Glukose an die Aminosäurenseitengruppe von Lysin oder Arginin unter Bildung einer Schiffschen Base. Nach einer Umlagerung entstehen sogenannte Amadoriprodukte. Bis zu diesem Punkt ist die chemische Reaktion prinzipiell umkehrbar. Das entstandene Amadoriprodukt ist für einige Wochen stabil. Das bekannteste Amadoriprodukt ist die modifizierte Form von Hämoglobin, HbA1c, das auch zur Diabeteskontrolle genutzt wird. Nach weiteren Umlagerungen, Oxidationen und Eliminationen entstehen die AGEs9 (Abb. 2). Die AGEs sind hinsichtlich ihrer chemischen Struktur sehr heterogen und können in toxische und nicht-toxische AGEs bzw. fluoreszierende und nicht fluoreszierende AGEs
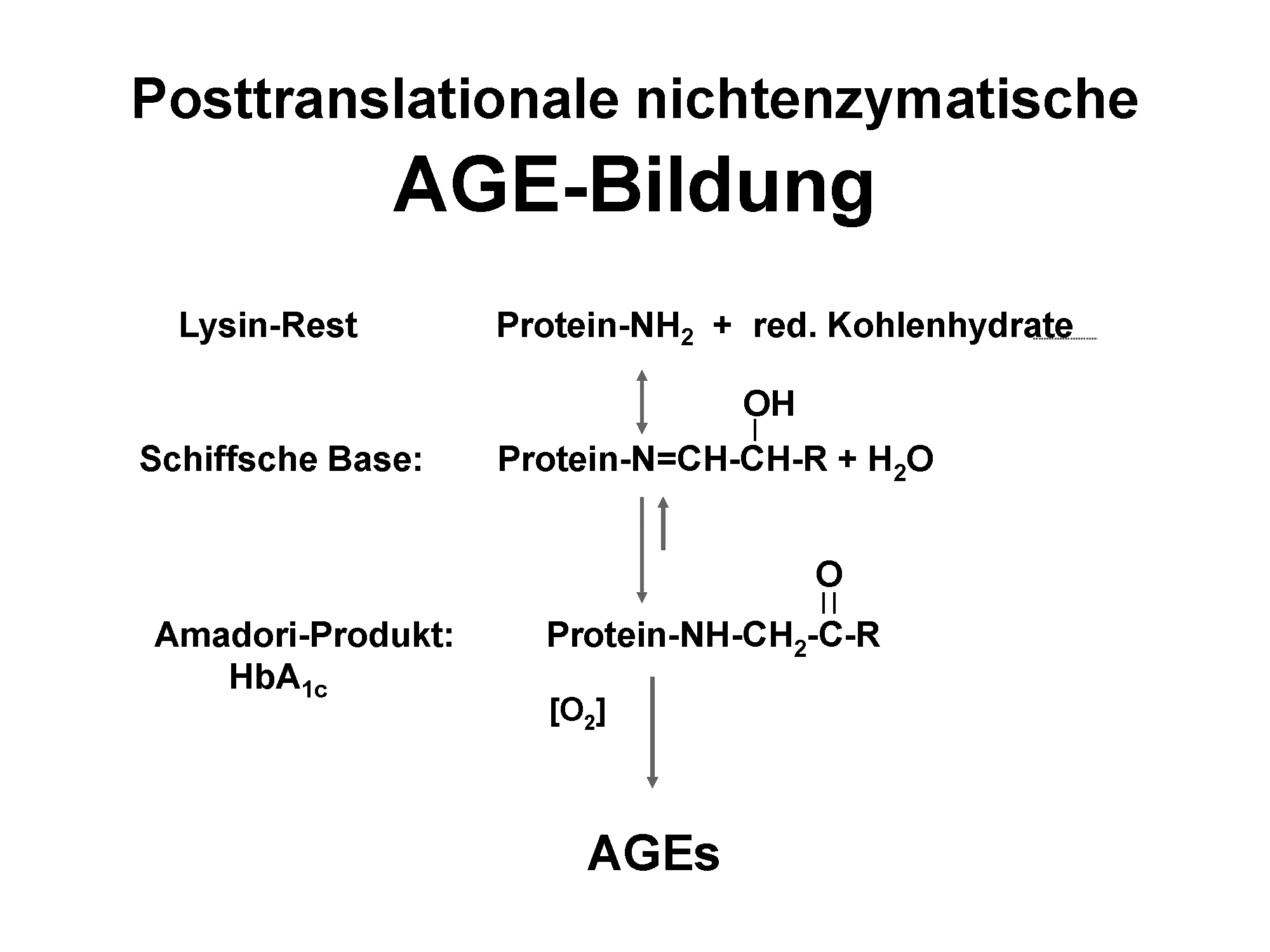 Abb. 2: Klassische AGE-Bildungsreaktion
Abb. 2: Klassische AGE-Bildungsreaktioneingeteilt werden.10 Zurzeit ist noch kein Enzym bekannt, das diese Modifikationen wieder vom Protein entfernen kann. Die einzige Möglichkeit, AGEs zu eliminieren, ist der Proteinabbau mit nachfolgender Ausscheidung der Abbauprodukte über die Niere. Jedoch ist der Abbau von Proteinen, die AGE-modifiziert sind, verlangsamt.11 Dies kann im Modell der Fruchtfliege zu einer Reduzierung der Lebensspanne führen.12 Die biologische Wirksamkeit dieser AGEs kann man grob in vier Bereiche einteilen: 1.) AGE-modifizierte Proteine verändern ihre Wirksamkeit, wobei sie zumeist an biologischer Aktivität verlieren. Dies konnte unsere Arbeitsgruppe am Beispiel des Wachstumsfaktors PDGF (Platelet-derived growth factor) und der nachfolgend verminderten Aktivierung von intrazellulären Proteinkinasen wie Akt oder ERK zeigen.13 Ein weiteres Beispiel stellt Albumin dar, das nach AGE-Modifikation eine reduzierte Esterase-Aktivität sowie geringere Bindungsaffinität an Ketoprofen aufweist.14 2.) Bei der AGE-Bildung kommt es häufig zur Quervernetzung von Proteinen, die Gewebsversteifungen und Proteinablagerung/Plaquebildung fördern. Dies ist insbesondere für das kardiovaskuläre System (Bluthochdruck, diastolische Herzinsuffizienz), die Altersweitsichtigkeit (Verlust der Elastizität der Augenlinse) oder die Alzheimer-Demenz (Alzheimer-Plaques) wichtig. Durch den modifikationsbedingt reduzierten Abbau von Proteinen wird natürlich auch der Proteinumsatz negativ beeinflusst, was wiederum zur Akkumulation langlebiger Proteine und somit Fibrose führt. 3.) Bei der Bildung von AGEs entstehen auch Radikale, die ihrerseits wieder Moleküle schädigen/oxidieren können.15 4.) AGEs können an Rezeptoren wie dem Rezeptor für AGEs (RAGE) binden und darüber die Expression proinflammatorischer Zytokine induzieren.16
Die Akkumulation der AGEs im Gewebe konnte schon vor vielen Jahren gezeigt werden.17 Da die oben genannten vier Wirkbereiche der AGEs primär auf experimentellen Versuchen beruhen, bleibt bisher eine zentrale Frage, ob AGEs tatsächlich eine pathophysiologische Relevanz beim Menschen haben. So konnte eine Studie mit Patienten, die sich einer Herzoperation unterziehen mussten, zeigen, dass der Gehalt an AGE-Modifikationen im isolierten quervernetzten Kollagen des Herzmuskels sowohl mit dem Alter des Patienten als auch mit dem negativen ›postoperativen Outcome‹ korreliert.18 Aus technischen und zeitlichen Gründen können solche aufwendigen Untersuchungen aber nicht in der klinischen Praxis umgesetzt werden. Um diese Problematik zu umgehen, können AGE-Ansammlungen auch im Kollagen der Haut nicht-invasiv mittels Fluoreszenzanalytik bestimmt werden, da deren Gehalt in der Haut mit denen des Myokards (Herzmuskelgewebes)19 und vermutlich anderer Organe vergleichbar ist. Eine zusätzliche physiologische Bedeutung bekommt dies dadurch, dass die Hautfluoreszenzwerte mit der Gefäßfunktion, die anhand der Pulswellengeschwindigkeit bestimmt worden ist, korrelieren.20 Damit spiegelt ein nicht-invasiver Analysetest primäre Alterungsvorgänge im kardiovaskulären System wider, was ihn als Biomarkertest für das Alter bzw. den alten Menschen interessant macht. Tatsächlich konnte eine Studie mit Diabetikern den Vorhersagewert dieser Fluoreszenzanalytik an der Haut zeigen. Patienten mit hohen Hautautofluoreszenzwerten (hohe AGE-Akkumulation) hatten eine 5-Jahres-Überlebensrate von unter 10 %, während die von Patienten mit geringen Werten bei 90 % lag.21
Die pathophysiologische Relevanz von AGEs über das AGE-RAGE System ist dagegen schwer in klinischen Studien zu ermitteln. Allerdings konnte in experimentellen Studien an Mäusen gezeigt werden, dass die Gabe von löslichem RAGE (sRAGE) – als antagonistischen Inhibitor der AGE-Bindung an RAGE – die Entstehung einer Diabetes-induzierten Arteriosklerose verhindert.22 Auch die Myokardinfarktexpansion ist durch blockierende Antikörper gegen RAGE reduziert, was durch Untersuchungen am RAGEknockout-Mausmodell bestätigt werden konnte.23 RAGEknockout-Mäuse sind zudem besser vor Sepsis geschützt.24 Bei Myokardinfarkt und Sepsis könnten allerdings auch andere RAGE-bindende Liganden als AGEs eine Rolle spielen.25 Unumstritten ist dagegen die RAGE-unabhängige Bedeutung von AGEs bei der zunehmenden Gewebsversteifung mit höherem Alter. So verbessern AGE-crosslinkbreaker wie Alt-711 wesentlich die Pumpleistung eines alten Herzens im Hundemodell,26 was deren nachteilige Bedeutung bei der kardiovaskulären Funktion unterstreicht.
Im Gegensatz zum menschlichen Organismus haben wir in erhitzter gebräunter Nahrung große Mengen an AGEs. Diese werden auch ins Blut aufgenommen und über die Niere wieder abgegeben.27 Ob die Nahrungsmittel-AGEs ebenfalls toxisch sind, wird zurzeit kontrovers diskutiert. So führte in einem Mausmodell eine AGE-arme Ernährung zu einer verlängerten Lebensspanne der Tiere,28 was deren mögliche Toxizität unterstreicht. Es gibt aber auch Hinweise, dass das nicht automatisch so sein muss. AGEs aus der Brotkruste scheinen zunächst auch einen negativen biologischen Effekt zu haben, da sie intrazellulär oxidativen Stress induzieren. Aufgrund der dadurch aktivierten Expression antioxidativer Enzyme wie den Superoxiddismutasen, führt dies in der Konsequenz aber dazu, dass so behandelte Zellen besser gegen oxidativen Stress und dadurch induzierten Zelltod geschützt sind.29 AGEs würden somit im Sinne einer hormetischen Antwort erst Stress induzieren und langfristig vor Stress schützen. Eine weitere unerwartete Ausnahme stellt das Bronchialkarzinom dar. Auch hier haben AGEs einen vermutlich positiven Effekt, da eine AGE-reiche Ernährung das induzierte Tumorwachstum am Nacktmausmodell verlangsamt.30 Klinische Studien an Patienten mit operablem Bronchialkarzinom konnten diese experimentelle Beobachtung bestätigen. So zeigten Bronchialkarzinom-Patienten mit hohen AGE-Gehalten im Blutplasma eine bessere 5-Jahres-Überlebensrate als diejenigen mit geringen AGE-Gehalten. Auch Diabetiker und insbesondere niereninsuffiziente Diabetiker, die generell hohe AGE-Gehalte im Blutplasma aufweisen, überlebten das Bronchialkarzinom besser als diejenigen ohne Diabetes und Niereninsuffizienz.31 Insgesamt zeigen diese Daten, dass hohe AGE-Gehalte in bestimmten Konstellationen auch protektiv sein können, Alternsprozesse also nicht immer nur negativ sein müssen. Zwei weitere Beispiele sollen diese gegensätzlichen Funktionen von Alterungsprozessen verdeutlichen. Das Altern auf zellulärer Ebene, d. h. der damit verbundene Replikationsstopp und die veränderte Zellfunktion, wird als replikative Seneszenz bezeichnet. Sie ist einerseits mitverantwortlich für die Gewebsinflammation im Alter (negativ), ist aber in jungen Jahren wichtig für die Tumorabwehr und bei der Wundheilung.32 Die familiäre Form der Hypercholesterinämie (erhöhter Cholesterinspiegel aufgrund eines Gendefekts) ist mit einer Frequenz von ca. 1 : 300 pro Geburt sehr häufig. Sie führt im hohen Alter zur Arteriosklerose, könnte aber in jungen Jahren protektiv bei Lungenentzündung sein.33
Kann man diese Alterungsprozesse selbst aktiv beeinflussen? Am leichtesten wäre es, wenn es einen Jungbrunnen gäbe, wie ihn Lucas Cranach d. Ä. (1472 bis 1553) in seinem berühmten Gemälde von 1546 dargestellt hat. Leider gibt es diesen nicht, und wenn doch, funktioniert er – in der Darstellung Cranachs – anscheinend nur bei Frauen. Die beste uns bekannte Möglichkeit, das Altern zu beeinflussen, ist körperliche Aktivität. In einer großen Studie konnte 2011 gezeigt werden, dass die allgemeine Mortalitätsrate mit dem wöchentlichen Aktivitätslevel sinkt.34 Bei älteren Menschen hat Tanzen einen positiven Effekt auf die kognitive Leistungsfähigkeit, gemessen an der fluiden Intelligenz und dem Arbeitsgedächtnis.35 Auch die Akkumulation der AGEs in der Haut lässt sich durch Sport positiv beeinflussen. Leider trifft dies aber nicht auf jeden zu.36 Dieser differentielle Effekt ist seit langem bekannt. In vielen Therapien, wie z. B. zur Behandlung von Bluthochdruck, gibt es nicht nachvollziehbare Non-responder, die auf eine medikamentöse Therapie nicht ansprechen. Warum präventive und therapeutischen Maßnahmen bei manchen Patienten wirksam sind und bei anderen eben nicht, wird wohl noch lange Gegenstand der Forschung sein.
Zusammenfassend kann man sagen, dass Alterungsprozesse pathophysiologisch bedeutsam sind und somit auch Basis von neuen Interventionsstrategien sein könnten. Andererseits gibt es aber Hinweise, dass dieselben Mechanismen in anderen Zusammenhängen protektiv sein können. Alterungsprozesse sind also nicht grundsätzlich negativ zu bewerten. Altern lässt sich auch positiv durch unseren Lebensstil beeinflussen, hier sind aber noch viele Studien nötig, um langfristig individualisierte therapeutische Ansätze zu identifizieren. Lassen Sie mich mit einem Spruch enden, der I. Bergmann zugeschrieben wird: »Älter werden ist, wie auf einen Berg steigen; je höher man kommt, umso mehr Kräfte sind verbraucht; aber umso weiter sieht man«.
- 1Richard G. Cutler,»Evolution of human longevity: a critical overview«, in Mechanisms of Ageing and Development 9/3–4 (1979), S. 337–354.
- 2Ebd.
- 3Jim Oeppen und James W. Vaupe, »Demography. Broken limits to life expectancy«, in Science 296/5570 (2002), S. 1029–1031.
- 4S. Jay Olshansky u. a., »A Potential Decline in Life Expectancy in the United States in the 21st Century«, in The New England Journal of Medicine 352/11 (2005), S. 1138–1145.
- 5Edward G. Lakatta und Daniel Levy, »Arterial and Cardiac Aging: Major Shareholders in Cardiovascular Disease Enterprises. Part I: Aging Arteries: A ›Set Up‹ for Vascular Disease«, in Circulation 107/1 (2003), S. 139–146.
- 6Thomas B. L. Kirkwood, »A systematic look at an old problem«, in Nature 451/7179 (2008), S. 644–647.
- 7Andreas Simm u. a., »Potential biomarkers of ageing«, in Biological Chemistry 389/3 (2008), S. 257–265.
- 8Louis Camille Maillard, »Action des acides aminés sur les sucres: formation des mélanoïdines par voie méthodique«, in Comptes rendus hebdomadaires des séances de l’Académie des Sciences 154/2 (1912), S. 66–68.
- 9Christiane Ott u. a., »Role of advanced glycation end products in cellular signaling«, in Redox Biology 2 (2014), S. 411–429.
- 10Takashi Sato u. a., »TAGE (Toxic AGEs) Theory in Diabetic Complications«, in Current Molecular Medicine 6/3 (2006), S. 351–358; ders. u. a., »Toxic Advanced Glycation End Products (TAGE) Theory in Alzheimer’s Disease«, in American Journal of Alzheimer’s Disease and Other Dementias 21/3 (2006), S. 197–208.
- 11Alexandra Stolzing u. a., »Degradation of glycated bovine serum albumin in microglial cells«, in Free Radical Biology and Medicine 40/6 (2006), S. 1017–1027.
- 12Eleni N. Tsakiri, u. a., »Diet-derived advanced glycation end products or lipofuscin disrupts proteostasis and reduces life span in ›Drosophila melanogaster‹«, in Free Radical Biology and Medicine 65 (2013), S. 1155–1163.
- 13Norbert Nass u. a., »Glycation of PDGF results in decreased biological activity«, in The International Journal of Biochemistry & Cell Biology 42/5 (2010), S. 749–754.
- 14Naila Ahmed, »Peptide Mapping Identifies Hotspot Site of Modification in Human Serum Albumin by Methylglyoxal Involved in Ligand Binding and Esterase Activity«, in The Journal of Biological Chemistry 280/7 (2005), S. 5724–5732.
- 15James V. Hunt, Roger T. Dean und Simon P. Wolff, »Hydroxyl radical production and autoxidative glycosylation. Glucose autoxidation as the cause of protein damage in the experimental glycation model of diabetes mellitus and ageing«, in Biochemical Journal 256/1 (1988). S. 205–212.
- 16Angelika Bierhaus u. a., »Advanced Glycation End Product-Induced Activation of NF-κB is Suppressed by α-Lipoic Acid in Cultured Endothelial Cells«, in Diabetes 46/9 (1997), S. 1481–1490.
- 17Erwin D. Schleicher, Eva Wagner und Andreas G. Nerlich, »Increased Accumulation of the Glycoxidation Product Nε-(carboxymethyl)lysine in Human Tissues in Diabetes and Aging«, in The Journal of Clinical Investigation 99/3 (1997), S. 457–468.
- 18Britt Hofmann u. a., »Relationship between cardiac tissue glycation and skin autofluorescence in patients with coronary artery disease«, in Diabetes & Metabolism, Onlinefassung v. 29. 12. 2014, http://www.diabet-metabolism.com/article/S1262-3636%2814%2900196-7/abstract (3. 11. 2015).
- 19Britt Hofmann u. a., »Advanced glycation end product associated skin autofluorescence: a mirror of vascular function?«, in Experimental Gerontology 48/1 (2013), S. 38–44.
- 20Ebd.
- 21Robbert Meerwaldt u. a., »Skin Autofluorescence is a Strong Predictor of Cardiac Mortality in Diabetes«, in Diabetes Care 30/1 2007, S. 107–112.
- 22Lisa Park u. a., »Suppression of accelerated diabetic atherosclerosis by the soluble receptor for advanced glycation endproducts«, in Nature Medicine 4/9 (1998), S. 1025–1031.
- 23Loredana G. Bucciarelli u. a., » RAGE and modulation of ischemic injury in the diabetic myocardium«, in Diabetes 57/7 (2008), S. 1941–1951.
- 24Birgit Liliensiek u. a., »Receptor for advanced glycation end products (RAGE) regulates sepsis but not the adaptive immune response«, in The Journal of Clinical Investigation 113/11 (2004), S. 1641–1650.
- 25Luis Ulloa und Davorka Messmer, »High-mobility group box 1 (HMGB1) protein: Friend and foe«, in Cytokine & Growth Factor Reviews 17/3 (2006), S. 189-201.
- 26Mohammad Asif u. a., »An advanced glycation endproduct cross-link breaker can reverse age-related increases in myocardial stiffness«, in Proceedings of the National Academy of Sciences of the United States of America 97/6 (2000), S. 2809–2813.
- 27Theodore Koschinsky u. a., »Orally absorbed reactive glycation products (glycotoxins): An environmental risk factor in diabetic nephropathy«, in Proceedings of the National Academy of Sciences of the United States of America 94/12 (1997), S. 6474–6479.
- 28Weijing Cai u. a., »Reduced oxidant stress and extended lifespan in mice exposed to a low glycotoxin diet: association with increased AGER1 expression«, in American Journal of Pathology 170/6 (2007), S. 1893–1902.
- 29Stefanie Ruhs u. a., »Preconditioning with Maillard reaction products improves antioxidant defence leading to increased stress tolerance in cardiac cells«, in Experimental Gerontology 45/10 (2010), S. 752–762.
- 30Babett Bartling u. a., »Prognostic Potential and Tumor Growth-Inhibiting Effect of Plasma Advanced Glycation End Products in Non-Small Cell Lung Carcinoma«, in Molecular Medicine 17/9–10 (2011), S. 980–989.
- 31Ebd.; Babett Bartling u. a., »Effect of Diabetes Mellitus on the Outcome of Patients with Resected Non-Small Cell Lung Carcinoma«, in Gerontology 57/6 (2011), S. 497–501.
- 32Marco Demaria u. a., »An Essential Role for Senescent Cells in Optimal Wound Healing through Secretion of PDGF-AA«, in Developmental Cell 31/6 (2014), S. 722–733.
- 33Martina Weber u. a., »Hepatic induction of cholesterol biosynthesis reflects a remote adaptive response to pneumococcal pneumonia«, in The FASEB Journal 26/6 (2012), S. 2424–2436.
- 34Chi Pang Wen u. a., »Minimum amount of physical activity for reduced mortality and extended life expectancy: a prospective cohort study«, in The Lancet 378/9798 (2011), S. 1244–1253.
- 35Kathrin Rehfeld u. a., »Auswirkungen einer Tanz- und Kraft-Ausdauer-Intervention auf kognitive Fähigkeiten älterer Menschen«, in Zeitschrift für Neuropsychologie 25 (2014), S. 99–108.
- 36Anne Navarrete Santos u. a., »Einfluss von Gesundheitssport auf Marker des biofunktionalen Alters: Nicht invasive Analyse von ›advanced glycation endproducts‹ in der Haut«, in Bewegungstherapie und Gesundheitssport 27 (2011), S. 121–124.